
日本人の祖先をDNAで探る:Y染色体ハプログループの分布
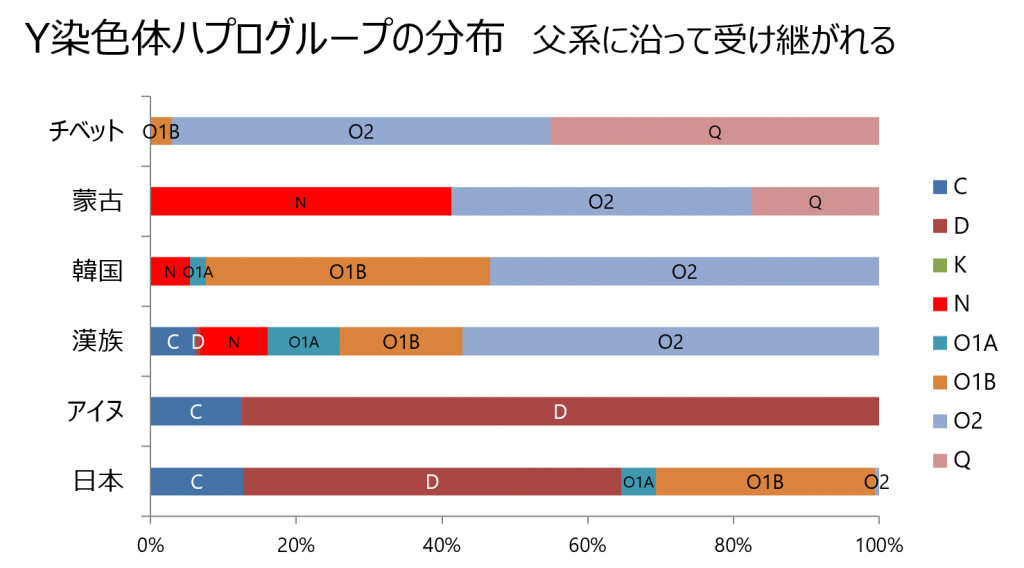
上の図は、Y染色体ハプログループの東アジアでの分布を示したグラフです。
Y染色体は男性のみに受け継がれるDNAです。
なので、為政者や戦争によって征服された状況を表しているともいえます。
この図からわかるように日本列島では、Y染色体ハプログループ分布の多様性が
高いことが明らかです。
ハプログループDは古代縄文人の末裔
ハプログループDは、現在の中国、朝鮮、東南アジアにおいて多数みられるO系統や
その他E系統以外のユーラシア系統(C,I,J,N,Rなど)とは分岐から7万年以上たっています。
また、世界で日本列島のみに見られるDが多く、日本列島固有のC1が少なからず見られます。
崎谷満氏は、現生人類は出アフリカ後、南ルート、北ルート、西ルートの3方向に拡散したとし
Y染色体ハプログープCの下位系統(の祖型)はそれぞれ分かれて別ルートを進んだ。
ハプログループDは、南北アメリカ大陸では全く見つかっていない。
D系統は東アジアにおける最古層のタイプと想定できるが、一つの説として東アジア及び
東南アジアにO系統が広く流入した為、島国日本や山岳チベットにのみD系統が残った。
そのため形質人類学的には古モンゴロイド(アイノイド)の分布と相関しているようである。
ハプログループC1a1
そして謎の多いのがC1a1です。
まだ学術的に解明されておらず、今後の研究が待たれるとされています。
C1a2が旧石器時代のヨーロッパ人であると言われており、その系統が枝分かれしたC1a1がなぜ日本で見つかるのか不思議です。
そして、C1a1が鳥取県の青谷上寺地遺跡で見つかった人骨に含まれていたとわかり、中国大陸を経由したと想定することも可能です。
ハプログループC1の下位群C1aのうち「M8, M105, M131, P122」によって定義されるグループである。
ハプログループC1a1 Wikipediaより
日本列島ではおおむね5%の頻度で発見されており、他には韓国の済州島で1人、中国遼寧省2人、中国河北省1人、北朝鮮の黄海南道1人、韓国3人が確認されている。
最も近縁なハプログループであるC1a2との最も近い共通祖先はおおよそ四、五万年前にさかのぼると推定されているが、(現段階の研究で把握されている限りでは)現存するものの拡散は約12,000年前と推定される。
現存する下位系統の最も近い共通祖先は日本列島で誕生したとも考えられるが、詳細は今後の研究が待たれる。
ハプログループC1a1(M8)の祖型の移動ルートは謎に包まれている。
鳥取県鳥取市青谷上寺地遺跡で発掘された弥生時代後期とみられる人骨について、県が国立科学博物館や国立歴史民俗博物館が分析した父系結果、4点のうち2点はハプログループC1a1(M8)であった。
二重構造モデル「縄文人」「弥生人」は嘘だった
朝鮮半島では、ハプログループD系統が見られるが(この図にはない)、近世にチベットから
モンゴル経由で入ってきたD1a1や、弥生時代に日本列島から朝鮮半島へ北上したD1a2aの系統
であろうと推測されている。
日本列島中間部でハプログループOは、約25%程度となる。
日本列島へはこの圧倒的な漢民族の膨張圧があまり及んでいないことを示している。
従って、東アジアとは異なり、日本列島ではハプログループO1以外のヒト集団が8割を占め、
さらに漢民族固有のO2が僅かしかいないことは、この日本列島が多様性維持の条件を
備えていて、東アジアの民族間の軋轢、抗争によって排除されたものも暖かく迎え入れて、この地
での存続を保証してきたことを想定させる。
二重構造モデルの問題点
- 多地域進化説を前提とする二重構造モデルでは「縄文人」「弥生人」ともそれぞれ別の人種、
別の生物学的亜種~種であるとする前提の制約を受けている。
分子人類学による単系統説、約20万年前にアフリカで生まれた単一種であることが実証されて
いるので二重構造モデルのよって立っている前提が崩れてしまった。 - 後期旧石器時代・新石器時代に日本列島へ流入したヒト集団は多様である。
「縄文人」が単一であるかのような見方は分子人類学的に否定された。
同時に完新世後半の水稲農耕開始期に流入したヒト集団も多様である。
「弥生人」が単一であるかのような見方も分子人類学的に否定された。 - 後期旧石器時代・新石器時代集団、水稲農耕開始期集団のいずれもが北方系である。
「縄文人」南方由来説は分子人類学的に否定された。 - 二重構造モデルによるアイヌ・琉球同祖論は、やや劣った別人種である「縄文人」が
北海道と琉球に残存したというものであったが、分子人類学的には誤っていることが
はっきりとした。アイヌ民族と琉球民族とはそれぞれ別の実体を持つ別集団である。 - 二重構造モデルでは、九州や西日本において「弥生人」によって「縄文人」が駆逐され
事実上ヒト集団の置き換えが起こったかのような説を示していた。
また100万人単位の「弥生人」の流入があったかのような説もあった。
いずれも、考古学的、形質人類学的、分子人類学的に否定された説である。 - 日本列島においては新石器時代に栽培農耕、雑穀農耕(ミレット農耕)などが開始
されていた。農耕は「弥生人」だけに独占されるものではない。
また、日本語も先住系集団に温存されていた言語に由来するものであり、新たに「弥生人」が
持ち込んだものではない。
その背景には、文化的なものはすべて中華文明に由来するという「中華文明絶対主義」
そしてその対である自己文化否定、「謙日本主義」のようなネガティブな側面が含まれるようである。
と言っています。
長浜浩明氏は、日本人男性の祖先の多くが”韓半島を通って大陸から波状的にやって来た”のなら
日本人のパターンは韓国人や中国人に似て然るべきです。
しかし、日本人は彼らと全く異なるパターンを持っており、それはD1Bにあることは明らかです。
と言っています。
